 Диэнай
Диэнай
После вспышки тяжелого острого респираторного синдрома коронавируса 2 (SARS-CoV-2), который является вирусом, вызывающим коронавирусное заболевание 2019 года (COVID-19), значительно увеличилось количество исследований, посвященных зоонозам.
Исследование: SDAV, крысиный коронавирус – сколько мы знаем о нем в свете потенциальных зоонозов. Изображение предоставлено: 22 августа / Shutterstock.com
Справочная информация
Некоторые из животных, которые были изучены, включают летучих мышей ( Rousettus aegyptiacus ), подковоносов ( Rhinolophus spp .), Рыжих полевок ( Myodes glareolus ]), енотов, собак, хорьков, норок, а также других домашних животных, таких как коровы и свиньи. Эти исследования в основном проводились для оценки того, экспрессируют ли эти животные рецептор ангиотензинпревращающего фермента 2 (ACE2), который является основным рецептором входа в клетки для SARS-CoV-2.
Хотя большинство этих исследований было сосредоточено на летучих мышах, следует уделять больше внимания грызунам, особенно крысам. Известно, что грызуны являются важными переносчиками или резервуарами некоторых зоонозных вирусов, таких как западный конский энцефалит, вирус лимфоцитарного хориоменингита (LCMV), геморрагическая лихорадка с почечным синдромом, болезнь, вызванная вирусом Апой, омская геморрагическая лихорадка и коровий гепатит E. Крысы также встречаются почти во всех частях земли; следовательно, люди с большей вероятностью будут взаимодействовать с этими животными по сравнению с любыми другими видами диких животных.
Новый обзор, опубликованный в журнале Viruses направлен на описание текущего уровня знаний о вирусе сиалодакриоаденита (SDAV), который является коронавирусом крыс, а также его характеристики и потенциальную зоонозную угрозу.
Классификация коронавирусов
Коронавирусы, как известно, принадлежат к отряду Nidovirales . Nidovirales характеризуются оболочечными, несегментированными РНК-вирусами с положительным смыслом.
Вирусы с наибольшей способностью вызывать пандемии и эпидемии относятся к подотряду Cornidovirineae порядка Nidovirales . Подсемейство Cornidovirineae Coronaviridae далее подразделяется на два семейства: Letovirinae и Orthocoronavirinae .
Orthocoronavirinae включает четыре рода коронавирусов: альфа, бета, гамма и дельта. Коронавирусы альфа и бета, как известно, заражают летучих мышей и млекопитающих, тогда как коронавирусы гамма и дельта заражают птиц и некоторых морских млекопитающих.
Обзор крысиных коронавирусов
Первое сообщение о коронавирусе крыс (RCoV) появилось в 1960 году после того, как у лабораторных крыс были обнаружены дакриоаденит, деструктивный сиаладенит и передающееся заболевание нижних дыхательных путей. После этого агенты, относящиеся к вирусу гепатита мышей (MHV), были обнаружены в сыворотке крови крыс в 1964 году.
Крысиный коронавирус Паркера (RCV-P) также был выделен из легких бессимптомных крыс. Известно, что второй штамм, вызывающий сиалоадакриоаденит (SDA), был антигенно идентичен RCV-P, также был распознан MHV. После этого в исследованиях оценивали несколько других штаммов RCoV.
RCoV относятся к бета-коронавирусам, особенно к подродам Embecovirus . Среди RCoV коронавирус China Rattus (ChRCoV) представляет мышиную линию Betacoronavirus 1 и их способность подвергаться межвидовой передаче от грызунов к другим млекопитающим. SDAV и другие RCoV принадлежат к тому же роду, что и человеческие коронавирусы, которые были способны вызывать пандемии и эпидемии во всем мире.
Передача SDAV, симптомы и диагноз
Экспериментальные исследования показали, что RCoV могут оставаться заразными при сушке на твердых поверхностях. Инфекции RCoV легко распространяются либо при прямом контакте с инфицированными людьми, либо через аэрозоль.
Известно, что RCoV вызывают симптоматические и бессимптомные инфекции. Также существует две модели заражения. Первая модель включает развитие гнездовых колоний, что в дальнейшем приводит к развитию эпизоотии у молодых неиммунных особей.
Вторая модель предполагает внезапное начало эписклерита у наивных крыс в период от отъема до взрослого возраста. Другие общие признаки инфекций SDAV включают отек подчелюстных слюнных желез, выделения из глаз, светобоязнь, слезотечение, язвы роговицы, помутнение роговицы и отек шейки матки из-за воспаления. Дополнительные побочные эффекты могут включать потерю веса, преходящую анорексию и нарушение течки.
Обнаружение SDAV и других RCoV достигается с помощью серологических тестов и иммуногистохимических методов. Полимеразная цепная реакция с обратной транскриптазой (ОТ-ПЦР) также используется для обнаружения генов M, N и pol, собранных из образцов инфицированной ткани. Лица с положительным диагнозом либо исключаются, либо помещаются в карантин на срок от 5 до 8 недель.
Структура SDAV и биологические функции белков
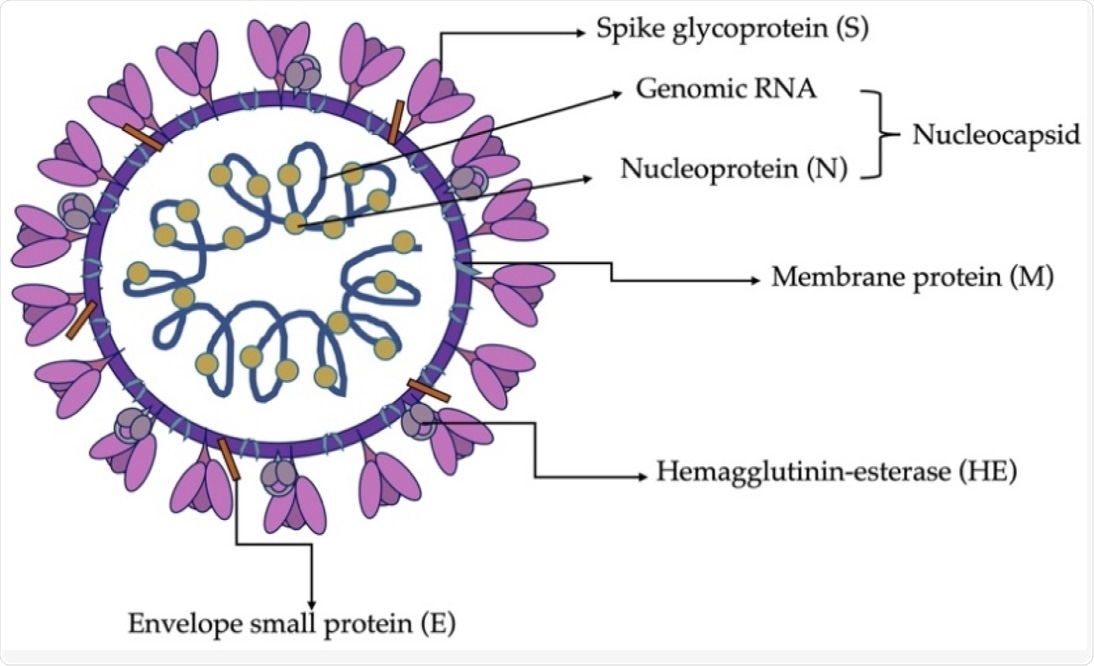 Схематическая структура вириона SDAV.
Схематическая структура вириона SDAV.
Вирион SDAV имеет сферическую форму диаметром от 80 до 180 нанометров (нм). Он включает ядро генома, окруженное мембранной оболочкой, и щупальца, похожие на шипованные белки, исходящие с поверхности. Вирусный геном состоит из четырех основных структурных белков, которые кодируются по направлению к 3'-концу генома, включая мембранные (M), шиповые (S), нуклеокапсидные (N) белки, а также гемагглютининэстеразу (HE).
Белок S отвечает за проникновение вириона в хозяйскую клетку. Области гена S-белка очень вариабельны, гетерогены и ответственны за тропизм тканей и изменение вирулентности.
Белок М отвечает за стимулирование кривизны мембраны за счет адаптации области мембраны для сборки вириона, а также за захват структурных остатков белка. Белок E работает в ассоциации с белком M во время сборки и морфогенеза вириона. Это также помогает в образовании пентамерных липидных пор в мембране хозяина, через которые может происходить перенос ионов.
N-белок способствует процессу репликации вируса и ограничивает трансляцию в клетках-хозяевах. Белки M, E и N работают вместе, вызывая сворачивание и почкование вновь собранного вириона. Роль белка HE заключается в прикреплении вириона, при этом он действует как поддерживающая связывающая молекула в дополнение к белку S.
Инфекция клеток и репликация вируса
SDAV способен инфицировать самые разные типы клеток, такие как эпителиальные клетки дыхательных путей, клетки центральной нервной системы (ЦНС) или мононуклеарные клетки лимфоидных органов. Точный рецептор, который способствует проникновению в клетки SDAV, еще не подтвержден. Предполагается, что SDAV использует механизм человеческого Betacoronavirus-1 и прикрепляется к рецептору сиаловой кислоты хозяина через их S-белок, чтобы способствовать проникновению в клетки.
После слияния с мембраной хозяина с помощью белков S и HE происходит протеолитическое расщепление белка S. При расщеплении белка S образуются две разные субъединицы, S1 и S2.
Известно, что расщепление происходит в двух разных точках: первая индуцируется фурином, а вторая – протеазой TMPRRS2, катепсином или любым другим белком. Это приводит к образованию слитого пептида S2 ’, который сливает хозяина и вирусную мембрану, таким образом вводя вирусную РНК в цитоплазму.
За этим следует трансляция гена репликазы, который помогает в репликации вируса. Трансляция гена репликазы приводит к образованию шестнадцати неструктурных белков (nsps), которые собираются в комплекс репликаза-транскриптаза (RTC) и осуществляют репликацию вируса.
Репликация включает геномную РНК с отрицательным смыслом, которая используется в качестве матрицы для синтеза положительной цепи. Репликация сопровождается транскрипцией, которая генерирует субгеномную положительную смысловую информационную РНК (мРНК).
Затем мРНК подвергается трансляции с образованием структурных и вспомогательных белков. Затем следует образование зрелых вирионов и их экзоцитоз. Точный механизм выхода SDAV еще не описан.
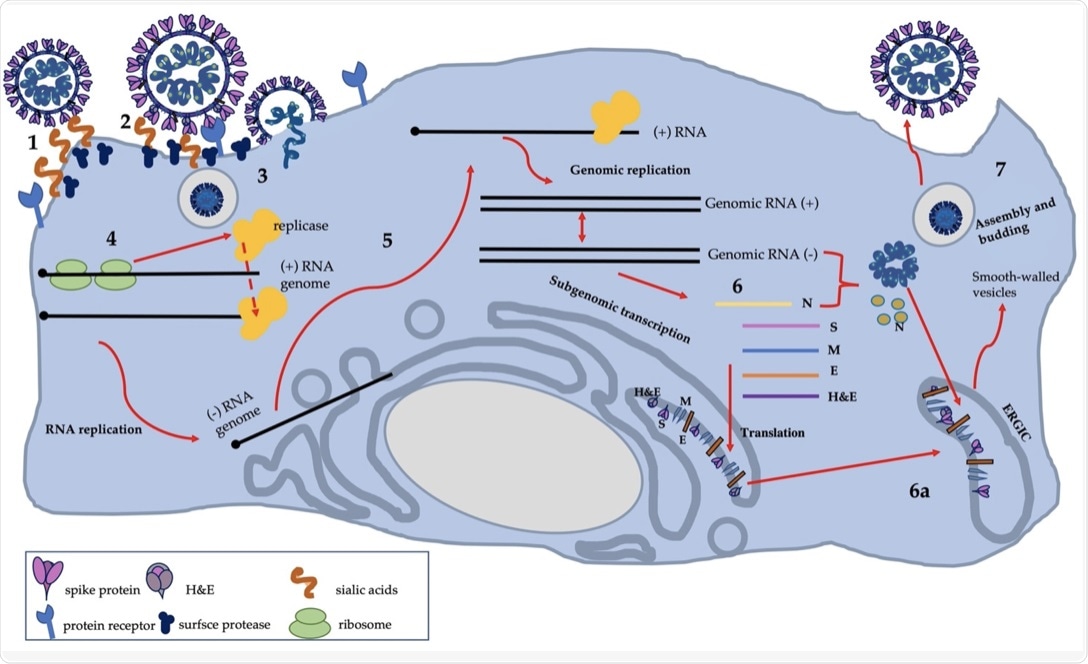 Схема репликации SDAV: собственная работа, основанная на имеющихся знаниях.
Схема репликации SDAV: собственная работа, основанная на имеющихся знаниях.
Геномная структура и функции генов SDAV
Характеристика геномной структуры SDAV была достигнута путем секвенирования 3’-концевых 9,8 т.п.н. геномной РНК. Семь фрагментов кДНК были получены путем клонирования комплементарной ДНК (кДНК) с помощью ОТ-ПЦР, которая представляла 3’-конец генома. Было подтверждено, что кодирующая последовательность для структурных белков S, M и N, а также неструктурных белков, NS2 и гена HE принадлежит девяти открытым рамкам считывания (ORF).
Кодирующая последовательность для ORF5a, неструктурного белка, M-белка и ORF7b, небольшой внутренней ORF, присутствовала в рамке +1. Кодирующая последовательность для полимеразы 1b, неструктурного белка 15k, малого мембранного белка и S-белка присутствовала в рамке +2.
Последовательности белка N, неструктурного белка NS2 и белка HE присутствовали в рамке +3. Кроме того, было обнаружено, что каждый из идентифицированных генов белка содержит короткую межгенную консенсусную последовательность.
Распространение SDAV и роль исследований in vitro
Ранее SDAV использовался для оценки ответа альвеолярных эпителиальных клеток на инфекцию. Недавно было установлено, что неинфицированные альвеолярные клетки I типа способны продуцировать хемокин в ответ на интерлейкин 1 (IL-1), продуцируемый SDAV-инфицированными клетками.
Продукция хемокинов наблюдалась также в клетках альвеолярного типа II и эпителиальных клетках дыхательных путей после инфицирования SDAV. Поэтому эпителиальные клетки альвеолярного типа I были обнаружены как первичная мишень для инфекции SDAV.
Текущие исследования также показывают, что размножение SDAV in vitro теперь возможно в различных установленных клеточных линиях, таких как линии клеток мышей и культуры клеток почек крыс, что упрощает экспериментальные исследования.
Заключение
Растущее число новых коронавирусов вызывает озабоченность из-за тесной связи коронавирусов с отдаленно родственными животными. Несмотря на сходство с зоонозными вирусами, зоонозный потенциал SDAV еще не исследован. Основной причиной этого может быть недостаточное знание всего цикла репликации вируса.
Симптоматическая инфекция SDAV легко поддается лечению, в то время как бессимптомные или легкие случаи трудно поддаются лечению и могут предоставить вирусу подходящую среду для мутации. Плохие гигиенические условия в клетках для крыс или мышей ответственны за передачу SDAV.
Хотя были описаны текущие исследования респираторных инфекций SDAV, необходимы дальнейшие исследования для идентификации взаимодействий вирус-клетка в различных клеточных моделях. Также необходимы дальнейшие исследования для характеристики инфекций SDAV и определения их зоонозного потенциала, который может представлять угрозу для человека в будущем.